The effort to increase food production levels and, as a result, food security, has led us to pen a number of articles about the advantages the C4 photosynthesis over plants and crops that use the C3 photosynthetic system.
In short, plants that use C4 photosynthesis have the ability to concentrate carbon around RuBisCo, thereby reducing the competition between oxygen and carbon atoms for this important enzyme’s binding site. Reducing the amount of oxygen able to be bound by RuBisCo reduces photorespiration levels, photorespiration being an energy intensive process that converts oxygen into CO2.
Given the superior carbon-fixing efficiency of C4 plants and resultant superiority in growth speed and efficiency, efforts have been focused by a number of researchers and research organisations on transplanting C4 photosynthesis systems into important crops that rely on C3 photosynthesis for carbon fixation and growth.
However, a recent article published in the Plant Biotechnology Journal looked more closely at the photorespiratory process of the C3 tobacco crop and, particularly, whether the process can be made more efficient to the benefit of plant growth.
Introduction
The researchers first note that previous research which limited or removed the photorespiration process resulted in reduced plant growth and development, even though the aim of those experiments was to remove the inefficiencies in carbon gain caused by the process. They then write about research that assisted the photorespiratory process by increasing flow through the glycine cleavage system (GCS) which, promisingly, resulted in increased biomass in Arabidopsis thaliana.
The GCS is described as a set of three enzymes (P, T and L) plus a small lipoylated protein (the H-protein) which moves between and interacts with the P, T and L enzymes. Previous experiments have shown that over-expression of the H-protein can result in an increase in GCS activity and biomass accumulation amongst other positive effects.
Following on from the previous research, the researchers looked at the effects of different promoters in front of the H-protein gene to increase expressiond on the growth of tobacco plants, using the constitutive promoter CaMV 35S and the light regulated ST-LS1 promoter.
Results
The researchers took an A. thaliana H-protein cDNA and over-expressed the gene in tobacco plants behind either the CaMV 35S promoter or the ST-LS1 promoter. Lines from each construct were self-fertilised to create T2 lines which were then subjected to quantitative reverse transcriptase PCR (qRT-PCR) and immunoblotting to quantify gene expression, confirm H-protein overexpression and identify homozygous and null mutant lines. Further, normal levels of expression of the L, P and T genes were confirmed.
H-protein expression levels showed that expression behind the CaMV promoter resulted in three to seven times more H-protein than when expressed behind the ST-LS1 promoter.
The effect of over-expression levels of the H-protein on plant growth were then assessed. The plant lines were grown in a controlled environment for 26 days after which leaf area and chlorophyll fluorescence levels were measured. The researchers found that the CaMV lines showed a significant reduction in leaf area compared to control plants (up to 50% less), while those lines where the H-protein was expressed behind the ST-LS1 promoter had leaf area similar to or up to 26% greater than control. No significant difference between chlorophyll fluorescence levels was found.
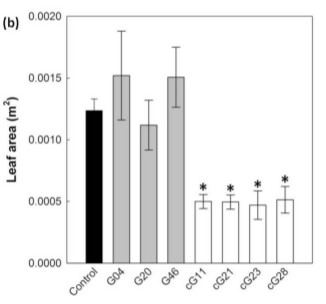
Figure 2b from article. Leaf area after 26 days of growth, demonstrating increase in leaf area in 2 of 3 ST-LS1 over-expression lines (in grey) and reduced leaf area in CaMV overexpression lines (white).
To analyse whether carbohydrate types and levels were a possible explanation behind the reduced leaf area, the researchers analysed carbohydrate profiles and found that whilst carbohydrate levels remained simlar between the CaMV lines and control, those transgenic lines had significantly lower levels of soluble sugars and increased starch accumulation. This alteration in profile was not seen in the ST-LS1 transgenic lines.
Lipoylation of the H-protein is essential to the function of the GCS, so the study looked at whether there was an alteration to lipoylation levels in the transgenic lines compared to control using immunoblotting of the lipoic acid groups in leaf and root tissue.What they found was that the lipoic acid levels in the transgenic plants were higher than in control. Whilst this makes sense given the higher H-protein levels in those lines, the greater levels of H-proteins in the CaMV lines compared to the ST-LS1 lines should result in a greater signal from the CaMV lines. However, the signal from both transgenic lines were similar, indicated that the extra H-proteins in the CaMV lines were not all active.
Given the negative effects of the CaMV promoter lines, the researchers chose to continue analysing the ST-LS1 plants further, testing the effects of the over-expression further under both glasshouse and field conditions.
Under glasshouse conditions, the transgenic lines showed increases in plant height, leaf number, leaf area and dry weight. Compared to control, above ground biomass of the transgenic lines were between 13% and 38% greater.
Glass house conditions are well and good, but the researchers wanted to see whether those increases were also possible under field conditions. So they performed two studies in consecutive years, the first being a small replicated control experiment using two transgenic lines which were grown for 25 days in a glasshouse before being moved into the field where they were assessed after 14 days. Once harvested, increases in biomass of both transgenic lines compared to control were again noted, resulting from increases in leaf number, height and total leaf area. Above ground biomass was 40% to 47% greater in transgenic lines compared to control.
The experiment was performed again the following year but this time using a larger, randomised block design. After being grown in the glasshouse for 33 days, the transgenic lines and controls were planted in the field and allowed to grow for a further 24 days and then harvested. Again, a significant increase in biomass was observed however one line, which had shown a significant increase over control in the previous year’s experiment, failed to outperform control. The other two transgenic ST-LS1 lines had increases in height (>18%), leaf area (>15%) and dry above ground biomass (>26%) compared to control.
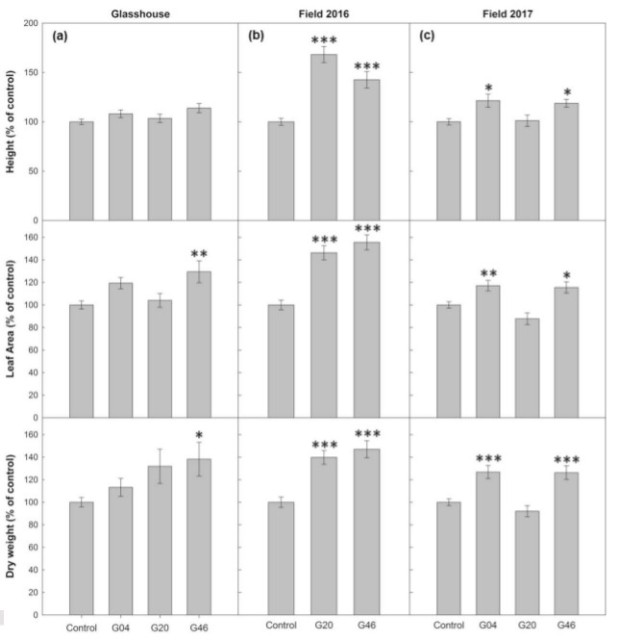
Figure 4 from article. Graphs depict differences in plant height, leaf area and dry weight between control and 3 ST-LS1 transgenic lines under greenhouse and two field experiements.
Testing whether increases in biomass were tied to changes in photosynthesis increases, the study analysed CO2 assimilation. They found that photosynthesis rates in ST-LS1 lines were increased compared to controls particularly under higher light intensities in glasshouse and field grown plants.
Finally, noting that conditions that trigger photorespiration are known to damage photosystem II, the researchers hypothesised that the increased photosynthesis rates in overexpressed H-protein lines may result from better protection of the photosystem from damage. To test the hypothesis, transgenic lines and controls were grown for nine days before being put in conditions of low O2 and continuous light – high photorespiratory conditions. They were subjected to these conditions for 24 hours before being returned to their original growth conditions and monitored for photosynthesis efficiency. The analysis found that photosynthesis efficiency was better preserved in transgenic lines compared to control.
Conclusion
The study finds that plants can have too much of a good thing when it comes to overexpression of the H-protein and GCS activity in tobacco plants. Whilst intermediate level of increased H-proteins resulting from gene promotion by the light-dependent ST-LS1 promoter resulted in increased biomass possibly due to improved photosynthesis rates and photosystem protection, too significant over-expression behind the constitutive CaMV failed for complete lipoylation of the extra H-protein, resulted in an altered carbohydrate profile and had reduced biomass compared to control plants. Importantly, growth under field conditions showed similar gains to those under controlled conditions.
The researchers have provided us with an alternative route to improving photosynthetic rates and crop growth rates than the more popular C4 engineering work. This study may provide either as an intermediate step towards greater crop production rates whilst C4 transformation research continues, be an improvement process that can employed in difficult-to-engineer plants or, should C4 transformation of C3 crops fail to become reality, improving photorespiration process could be one of many improvements in crop growth we see developed further in the future.